Copyright © Physiologia Plantarum 1999 (ISSN 00319317)
Dispersion Of Energies In Worldwide Crop Formations
Author: W. C. Levengooda and Nancy P. Talbottb
aPinelandia Biophysical Laboratory, 4853 Wolf Lake Rd., Grass Lake, Ml 49240, USA
bTalbott, BLT 0ffice, Box 400127, Cambridge, MA 02140, USA
Abstract:
The findings reported here lend further support to a decade of research which suggests that over 95% of worldwide crop formations involve organized ion plasma vortices that deliver lower atmosphere energy components of sufficient magnitude to produce significant bending, expansion and the formation of unique expulsion cavities in plant stem pulvini, as well as significant changes in seedling development. Here we demonstrate that, in a number of formations, quantitative node alterations can be directly related to fundamental concepts of electromagnetic energy absorption through the atmosphere. Also, in a detailed field study of simulated crop formations, we show that overfertilization or gravitropism cannot account for observed physiological alterations. In addition, we demon strate that details of flattened, intertwined crop patterns can be described by the application of fluid dynamic principles governing the interaction of rectilinearly moving vortex pairs.
Introduction
New findings reported here augment an ongoing investigation initiated in the late 1980s, when samples taken from 'crop formations' in wheat fields in southern England were first sent to the laboratory. Crop formations are flattened, swirled areas of plants, most often found in fields of commercially grown cereal plants such as wheat (Triticum aestivum), barley (Hordeum vulgare) or oats (Avena sativa); these flattened areas of the crop are often interspersed with areas of standing, apparently unaffected plants, the overall effect often being recognizable geometric patterns of considerable intricacy and dramatic effect.
Although simple flattened circles have been reported occasionally in southern England since the 1600s, by the mid1960s they were increasing in number, and by the late 1980s, not only numbered in the hundreds each summer, but had reached a level of design complexity which included not only circles and rings, but triangular and rectangular areas with associated pathways, often reaching 1-125 m in length or diameter. Most of these formations exhibited a number of features which had the farmers and local landowners mystified as to their causation.
During the 1993 growing season the British 'news' media initiated a story, subsequently circulated worldwide, that crop formations were the product of vandals. This assumption that all crop formations were manmade was not only accepted by much of the general public, but also, apparently, by the majority of scientists - in spite of the fact that there was absolutely no objective evidence presented to support such an idea. In fact, quite the opposite situation prevailed: although not yet published in 1993, considerable data had been analyzed in the laboratory, which conceptually offered a more sound and scientifically rigorous explanation.
By 1994 over 80 crop formations had been sampled from the USA, Canada, Australia and England and these plant specimens had been examined and analyzed in detail at the Pinelandia Laboratory at Grass Lake, Michigan, USA. By the time these results were published in the scientific literature (Levengood 1994) it was clear that the plants from more than 95% of the sampled events revealed either single or multiple anomalous and readily apparent structural alterations. In general, these consisted of significant enlargements in the internal cellwallpit structures, expanded and deformed stem pulvini, and gross embryonic malformations.
Not one of these clearly anomalous plant alterations had been mentioned - much less explained - by the proponents of the vandal theory, nor can they be accounted for by the supposed methods employed to create crop formations through claims made by the self described vandals.
One remarkable feature of plants within crop formations which, upon laboratory examination, displayed retarded or completely terminated embryo or seed development was the fact that the somatic tissue and external appearance of the plants were normal, thus producing a situation which can not be explained by ionizing radiation or chemicals - both known to alter development within somatic tissues (Leopold 1964, Levitt 1980). In addition, seeds taken from flattened plants in the formations regularly disclosed either increased or decreased seedling growth. These changes were closely associated with the particular growth stage at the time the crop formation energies became evident in the field. At the present time more than 250 crop formations have been examined, in a growing list of countries, as the phenomenon becomes more widely recognized and reported.
Not only does the research reported here verify earlier observations, it also presents quantitative evidence supporting the hypothesis of the involvement of complex plasma vortex energies as the causative process behind crop formations.
Materials and methods
Plant material
Procedures for collecting plant material from crop formations have focussed on obtaining sample sizes sufficient for statistical analyses and the sampling of plant tissue at a number of sites both within and outside the flattened areas. Because the vast majority of the crop formations examined in the laboratory have been in cereal grain crops, these species will be referred to when discussing the stem node analyses. At a given sampling site within a formation area or at a control location, a group of 15-20 plants are excised at around ground level. The field workers give each sample set a simple code number and carefully designate its location on a diagram of the formation. If the plants are moist and not at harvest maturity, they are dried down before shipment; this is of particular importance in the case of plants taken in foreign countries where shipping times can be from 10 to 14 days, thus allowing the buildup of severe mold in plants which have not been properly dried.
Analysis of stem pulvini
The most reliable and consistent node length data are obtained from apical and penultimate pulvini, with the direction of measurement along the vertical or longitudinal axis of the stem. The length of each node in a sample set is determined by using a microscope with low magnification or a 6 x loupe. A stem section (3-4 cm in length) containing the node region is placed beside a metal machinists scale graduated in 0.5 mm intervals. The length, estimated to the nearest 0.1 mm, is referenced from the dark lateral lines of tissue demarcation located at the basipetal and acropetal boundaries of the pulvinus region.
Method of examining gravitropic responses
The natural gravitropic responses in plant stem pulvini were examined by preparing carefully engineered, manmade crop circles in winter wheat grown in Maryland, USA, during the 1997 growing season. In addition to examining gravitropism, the authors were also interested in the influence of overfertilization on the stem nodes and other possible changes in the plants. This stem node study, begun on 5 May 1997, was carried out in commercially grown wheat in which two strips (3.7 x 122 m long) were staked out in a N-S direction, with a 3.7 m separation between. One test strip (strip A) was given twice the normal amount of fertilizer (pelleted urea, 46% nitrogen); the second strip (strip B) was treated with the normal amount of fertilizer (same as rest of the field grown for commercial harvest).
On 3 June 1997, after sampling upright normal 'base line' plants, two circles (1 and 3 m indiameter) were formed at the southern ends of both strips A and B. These four circles were formed using a plank with ropes attached at the ends (consistent with the proposed method used by selfproclaimed vandals) and then applying full body weight to the board as it was moved around in a circular motion for three or four rotations, thus forcing the plants down to create simulated crop circles. Additional identical pairs of circles were then created on 13 June 1997 at locations about 30 m north of the first circles, with a third set created on 25 June 1997, just prior to harvest, at a further distance of another 30 m.
At various intervals after the creation of each circle pair, 2-4 sample sets were collected from all flattened circles and from upright plants 3-76 m from the flattened areas. The node lengths (NL ) were recorded from each plant in a 15-20 plant sample set and routine statistical analyses (standard t-test; P<0.05 criteria) performed. The first two sets of test circles were formed when the plants were in rapid development and growth and an obvious gravitropic response, consisting of upward bending plants, was noted 48-72 h after the circle sets were created.
Results and discussion
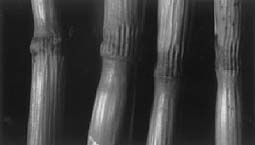
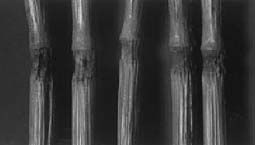
The most consistent alteration in cereal crop formations takes place within anatomical structures known as stem pulvini or growth nodes. In the majority of cases, the nodes on the formation plants are expanded relative to the normal, upright control plants taken from 0.3 to over 100 m outside the flattened areas. An important point to be emphasized is the fact that, within many formations, tufts of standing plants, generally located near the epicenter, have node expansions equal to, or often exceeding, the expansion level in the flattened plants. Analysis of stem nodes thus provides a convenient and quantitatively useful indicator of the degree of alteration or damage produced by the vortex energies. In addition to the length factor, the nodes are also examined for the presence of what the authors have defined as 'expulsion cavities', an anatomical alteration (shown in Fig. 1) which has never been observed in literally thousands of control plants examined in this laboratory. In a given formation, the frequency of expulsion cavities, when present, is consistently higher in the penultimate nodes compared with the apical pulvini. To understand the significance of these differences, the general mechanism of expulsion cavity formation must first be considered. Evidence has accumulated during the period of study which strongly suggests that the crop formation energies produce a very rapid and intense heating within the plant tissue (Levengood 1994). When this internal heating is of sufficient magnitude, pressure buildup within the node parenchyma tissue reaches the point where cellular components are literally blown out through the epidermal cell walls.
The viscoelastic nature of plant cell walls has been demonstrated in a quantitative manner (Ortega 1990). In those nodes exhibiting expulsion cavities the internal pressure produced a tension stress on the cell wall fibers, at a level which exceeded their elastic limit. At the apical node the tissue is less mature than the tissue at the penultimate position and, consequently, is more viscoelastic, or more easily expanded. This greater degree of apical node expansion capability reduces the internal pressure, whereas in the more mature and elastically rigid penultimate nodes the pressure is relieved by tissue rupture. In other words, the operation of two different types of pressure release 'valves' in the apical pulvini region is observed.
Gravitropic responses in plant stem pulvini
In control studies where maturing cereal grain plants are placed in a horizontal position a process of vertical recovery is initiated, the effects of which can be observed in just a few hours after the positioning (excellent time lapse photograph pp. 433 in Salisbury and Ross 1992). This gravitropic response is generally believed to be associated with the vertical movement of auxin (IAA) to the lower sides of the stems where it locally increases cell growth rates, thus producing an upward bending of the horizontal plant stems (Esau 1967, Wilkins 1984, Salisbury and Ross 1992). At the stem growth nodes this gravitropism effect produces a bending and lengthening of the pulvinus region which, in the case of crop formation plants, would be independent of the external energies.
 |
 |
| Fig. 1. Penultimate nodes on normal, upright control plants (A) and typical 'expulsion cavities' (B), in penultimate nodes from plants within the flattened area. Wheat crop formation at Logan, UT, USA, Aug.'96. | |
 |
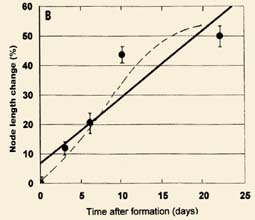 |
Fig. 2. Gravitropic influence on apical node lengths in flattened, manmade crop circles in wheat. Upper data set (A) from test strip given twice the normal fertilizer and the lower data set (B) from a test strip given the normal amount of fertilizer. Each point is the percentage node length change relative to normal, upright controls. Sample sets contain 15-20 plants each (vertical bars indicate SD). Solid lines, linear regression; dashed lines, demonstrate the sigmoid shape typical of growing tissue. Data taken between 3 and 25 June 1997. |
|
In studies of the stem node lengths as related to the crop formation energies, it became important to quantitatively define the role of gravitropism in the flattened formations. Although there is considerable literature relating to phototropism and gravitropism in A6ena coleoptiles, a paucity of information dealing with stem node responses under normal field conditions exists. In summarizing the node length variations from the previously described control study, the data from the 1 and 3 m circles in each set have been combined, because, within corresponding sample sets, there was no significant difference between the NL means. The results in Fig. 2 illustrate the node length changes in the strip A and strip B circles relative to the normal upright 'control' plants. Each point represents the mean node length change in a total population of 60-80 plants, with the standard deviations (SD) indicated by the vertical bars. The dashed lines in Fig. 2 indicate the sigmoid type of growth curve which is a ubiquitous characteristic of tissues in the cell wall extension growth phase (Salisbury and Ross 1992). The linear regression analyses (solid lines) have essentially identical slope constants, a clear indication that overfertilization does not influence the gravitropic response.
The plants taken 3 days after forming the circles have a mean node length increase of about 10% in both test strips. Within the data base it is observed that the majority of crop formations discovered in the field have been sampled within a time frame of a few hours to 3 days after their arrival and, as will be discussed later, the node length changes in these formations range from a 30 to an over 200% increase in node length relative to their upright controls. In these cases the gravitropic responses would therefore have very little influence on the overall node expansion analyses.
Within sample sets taken 3 days after forming the second circle pair (16 June 1997), the gravitropic responses were very comparable to those shown in Fig. 2, with ÿ+9.6% in strip A (n = 86) and +11.8% in strip B (n = 85). These very similar 3 day responses in two sample sets formed about 2 weeks apart suggest that the gravitropic responses remain fairly constant over the early stages of plant development.
Delineation of energies producing crop formations
The most basic attributes of the anomalous transformations taking place in the apical stem pulvini of crop formation plants are highly suggestive of an efficiently delivered energy which produces a very rapid heating within the stem node tissue. The question then becomes: how does one delineate this energy in a quantitative manner?
During the 1994 growing season, field workers sampled a number of crop formations with relatively simple circular geometric patterning, in which it was observed that the node damage decreased from the center to the edges of the flattened area with node expansion levels demonstrating a nonlinear trend with radial distance. If the energy causing node expansion is electromagnetic in origin (Levengood 1994), then the quantitative alterations in the plant stem nodes should fall within the framework of a straightforward and widely applied concept of physics dealing with the absorption of electromagnetic energy by matter, namely the BeerLambert principle (Moore 1962). As applied here, the fraction of the total energy striking a plant at a distance d (m) from a radiation source of intensity I0 ('point source' relative to the flattened area) is expressed by:
| I/I0 = e -ac(d) | (1) |
where I is the intensity of radiation at the plant, a, the molar absorption coefficient (m3 mol -1 m -1) of the intervening media and c, the molar concentration (mol m -3 )of absorbing molecules (air and water vapor). It is assumed that the stem node length, NL is directly related to the fraction of the energy absorbed into the node tissue, that is,
| NL = b ( I/I0 ) | (2) |
where b is the proportionality constant. From relationship (1) we now have:
| NL = b [ e -ac(d) ] | (3) |
and taking the logarithm of both sides of Equation (3) gives the useful expression:
| ln(NL ) = -ac(d) +b' | (4) |
where b' is the intercept constant. Since a and c are also constant, we would predict from relationship (4) a linear correlation between the log of node length and the distance d between the plants and the radiation source.
The node length data in Fig. 3 were obtained from a 1993 circular formation in a wheat field at Devises, England. Each point represents the mean node length (vertical bars indicate SD) in sample sets consisting of 10-15 plants per group. The horizontal dashed line displays the combined mean from four sets of controls (n = 40) taken at 7.6 to over 46 m from the formation. The linear regression curve exhibits excellent agreement (r = 0.91) between the empirical data and Equation (4) which is based on the electromagnetic energy absorption model.
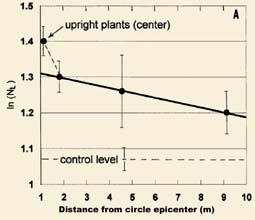
In many formations, circular shaped epicenter tufts of standing plants occur, which appear normal, ranging from 0.2 to 0.6 m in diameter. When the NL means from the standing epicenter tufts are compared with those obtained from samples taken from the flattened plants, one often finds that the ln(NL ) value from the epicenter lies well outside and above the regression curve for those in the flattened areas. Results from an 18 m diameter circle in a 1994 wheat field at Chehalis, Washington, USA, are presented in Fig. 4A. The data are presented as in Fig. 3 (n = 39 controls plants), with the solid line giving the linear regression (r = 0.99) for the downed plants.
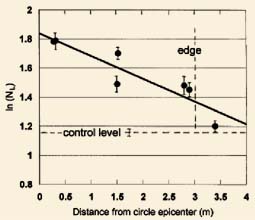 |
Fig. 3. Linear regression (r = 0.91) from crop formation data plotted according to the BeerLambert relationship dealing with the absorption of electromagnetic energy by matter (see text). Formation in a 1993 wheat field at Devises, England.
|
 |
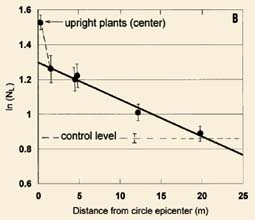 |
| Fig. 4. Very high node expansion levels in standing tufts of plants at the epicenter of crop formations (data plotted same as in Fig. 3). (A) 1994 wheat field at Chehalis, Washington and (B) 1994 barley field at Sussex, England. | |
A very similar standingplant epicenter effect is shown in Fig. 4B, in barley plants from a 1994 formation at Sussex, England. In this case the formation was a 'tear drop' shape. The linear regression (r = 0.98) includes two data points from plants taken at 12 and 20 m from the epicenter. The point at 12 m (or 6 m outside the formation) clearly displays significant node expansion, and is typical of many formations in which an energy 'spillover' effect is observed. This simply means that the energy producing the node expansion is not, in every case, totally confined within the visibly flattened area. At a distance of 20 m from the epicenter the node expansion is at the same level as in the controls taken 30-60 m from the event.
To explain the greater degree of node expansion or tissue damage in the standing plants (both within and sometimes outside the formation) the angle at which the radiation source impinges on the upright stems must be considered, and compared with the angle at which it strikes the flattened plants. Maximum energy transfer occurs when the surface being irradiated is orthogonal to the radiation source. If, in the upright plants the angle of incidence approaches 90°, then at the flattened stems the incident radiation would be at a more acute angle, resulting in less energy received unit area -1 . The fact that the node regions are rapidly heated internally, to the point of producing expulsion cavities, suggests a microwave type of energy source. The internode regions of the plant stems are tubular, whereas the pulvini are fully hydrated with a solid tissue crosssection, as a consequence, the pulvini have a higher dielectric constant and would receive a higher degree of microwave heating compared with the internode region.
Vortex interactions within crop formations
The wheat stem nodes shown in Fig. 5 are from a 1995 crop formation in Blue Ball, MD, and depict one of the most severe examples of pulvinus expansion examined in the laboratory. A total of ten sample sets (15-20 plants set -1 ) taken within a circular region about 27 m in diameter, disclosed node length increases ranging from 130 to over 200% relative to control plants (n = 95) taken outside the formation. This high level of expansion was extended throughout the formation and was most evident in the apical nodes. In the penultimate nodes 65-85% of the formation plants contained expulsion cavities (none were found in the controls). These expulsion cavities are, as previously pointed out, another indication of high energies, delivered very rapidly.
In addition to the node alterations this formation contained welldelineated, linear paths forming a raised ridge of plants (Fig. 6A) snaking in a sinusoidal pattern across the floor of the formation. Because of their unusual, linear extension, this ridge effect prompted the authors to examine the possible mechanisms of formation. From earlier work (Levengood 1994) the findings clearly suggested that the organization of a crop formation arises from instability products within selforganized ion plasma vortices.
 |
 |
| Fig. 5. Normal wheat nodes in upright controls (A), and severe node expansion (B) in flattened plants within a circular, 1995 crop formation at Blue Ball, MD, USA. | |
 |
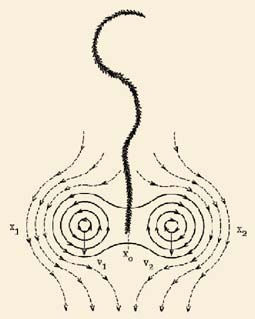 |
| Fig. 6. Ridge formation (A) within the flattened plant area of the circular crop formation at Blue Ball, MD, USA. Diagram (B) proposes, counterrotating vortices which produced plant 'ridges' in region X 0 of active reverse flow (see text). | |
From the familiar vortex appearance of the pattern in plants surrounding these ridges it seemed reasonable to apply fundamental fluid dynamic theory and examine the ridge pattern from the standpoint of interacting rectilinear, moving, vortices (Prandtl and Tietjens 1957, Peratt 1992). If a simple vortex filament aligned normal to the crop surface and having a core of constant rotation is considered, the velocity v1 on the circumference is connected with the angular velocity q, as:
|
v1 = q r1 |
(5) |
where r1 is the radius of the vortex core. Outside this core a potential plasma flow exists, within which the velocity v2 of movement at a distance r2 from the core center is:
|
v2 = T / 2p r2 |
(6) |
where T is the circulation strength or 'vorticity', and is given by:
|
T = 2q (p r12) |
(7) |
If a nearby vortex core approaches this vortex filament, its motion will be influenced by that of the neighboring system. If there are two mutually interacting vortices the potential fields or velocity distributions may become superposable. In the case of two vortices rotating in a parallel direction, and both with axes normal to the crop surface, the movements of their cores would have opposing velocity vectors, that is, their hydrodynamic interactions would cause the cores to move apart and in opposite directions. In this type of situation their mutual interactions would not form ridges of piled plants.
But, if the case of two interacting vortices with opposing rotations is examined, the core movements V1 and V2 are, as shown in Fig. 6B, in the same direction, and form a closed system around the individual vortex cores and an open system around the mutually interacting core pair. At the region X0 , midway between the vortices, the velocity is 1.5 times that in the open system X1 and X2 , and is in the opposite direction to the core movement. Thus the ridges are formed in the region of very active reverse flow. The interior plasmas accompany the vortices in their rectilinear movement through the crop, whereas the surrounding air flows around them as if they were a rigid body system. A reduced pressure occurs in the X0 region and this, combined with the high velocity profiles, would account for the plants being drawn upward and forming a ridge of piledup plants behind the vortex cores.
Intricacies of geometric design
Although many crop formations with very irregular outlines have been attributed to wind damage or overfertilization, we have determined that the same type of complex energy systems appears to be involved in their creation as in those displaying more geometric patterns. In fact plant damage of greater magnitudes and of more complex distributions occurs within the more irregularly outlined crop formations.One such formation with a roughly rectangular outline (38 m [E-W] x 50 m [N-S]) was sampled in a wheat field at Blue Ball, MD, at a site located approximately 1 mile from the circular formation previously discussed (both formations occurred on 29 May 1995)
At the east end were three 1.8 m wide paths of alternating standing and flattened plants. Within nine sample sets taken within the formation the node lengths ranged from a 92 to a 203% increase relative to the controls taken outside the formation, with the highest values occurring at the east end of the field at the regions of the alternating flattened:upright 1.8 m strips. When examining the energy absorption relationship (Equation (4)), there appeared to be a roughly decreasing trend, but the linear regression correlations were considerably reduced as compared with the ln(NL ) data from the sharply outlined, circular formations (such as those shown in Figs 3 and 4).
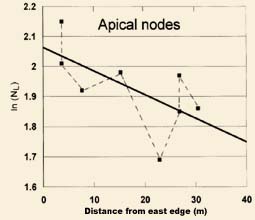
In Fig. 7 the data forming the upper regression curve (r = 0.64) were obtained from the apical nodes and the lower regression curve (r = 0.81) from the penultimate nodes. The dashed lines simply emphasize the deviation of the individual sample means, and it is of interest to note that the deviation pattern within the apical samples is almost identical in shape to that obtained from the penultimate nodes. These similar scatter patterns in node length data obtained at different positions on the plant indicate that the deviations from the regression curve are an inherent aspect of formation energy variations at the sampling sites, and are not measurement errors.
The data in Fig. 7 also indicate that more than one energy source was involved in producing the node expansions within this roughly rectangular formation.
 |
 |
| Fig. 7. Comparison of node length variations in apical nodes (upper regression curve) with variations in penultimate nodes (lower regression curve) from sample sets taken within a roughly rectangular outlined crop formation in a 1995 wheat field at Blue Ball, MD, USA. Note similar patterns (dashed lines) of sample set deviations from the regression lines, even though the node data were from different locations on the plant stems. | |
Table 1.
Putative energy components involved in the establishment of crop formations.
|
(I) Properties: |
||
|
(1) Ion plasma - self organized through chaos mechanisms |
||
|
(2) Dissipative system - far from equilibrium |
||
|
(3) Sensitive to initial boundary conditions |
||
|
(II) Composition of independently operating boundary conditions: |
||
|
(1) Microwaves - produce transient heating, node expansion and expulsion cavities in plant stem nodes |
||
|
(2) Electric fields produced by charge separation in plasma vortices-influence |
||
|
(3) Convective stability - motion of gasses inside vortex |
||
|
(4) Convective turbulence - may occur internally or from external weather conditions |
||
|
(5) Influence of earth's natural electric field locally this can vary from 1 to 100 V cm -1 - |
||
|
(6) Magnetic fields: |
||
|
(a) Produced by motion of ions and electrons inside plasma |
||
|
(b) Variations in earth's magnetic field (local) |
||
|
(7) Thermal gradients - inside and outside vortices |
||
The primary vortex originated at the east end, however, with conditions as shown in Fig. 6, other interacting vortices of lower energy content could locally perturb the primary source to the extent of producing significant local variations in the distributed energy. Within vortex rings produced in liquids it has been demonstrated (Levengood 1958, Rivest 1992) that very subtle, external convective turbulence can significantly change a welldefined, geometrically organized, vortex system into one with complete asymmetry.
Recently we have questioned why there is a greater percentage of reported (and subsequently sampled) crop formations in the USA which have highly irregular, nongeometric outlines, as compared with the preponderance of geometrically patterned events reported in England and several other countries. Here two factors should be considered: the human curiosity response related to the discovery of recognizable geometric patterns; and the basic dynamics of the forces involved in crop formation development.
Areas of nongeometric, randomly flattened cereal crops have apparently been observed by farmers worldwide for a considerable period of time and have been casually attributed to weather damage, or to overfertilization with subsequent weather damage, or even to animal and bird intervention. Such randomly flattened areas have excited little investigation and have been essentially ignored. When more geometric areas of flattened crop began to be observed with increasing frequency it was thought that this was an entirely separate, unrelated phenomenon. And, because the majority of the reporters of these more geometric events were primarily nonscientists, unaware that a relationship might exist, the irregularly flattened crop has been largely ignored as having mundane explanations. This is still primarily the case today. However, aerial photographs of crop formations in every country where they are currently reported, frequently reveal areas of irregularly flattened crops in conjunction with geometrically flattened patterns - either in the same, or adjacent fields. It is simply that the geometric designs excite much more interest.
Also, the basic dynamics of the forces involved in crop formation development are not immediately apparent. In any complex phenomenon in which chaotic mechanisms are implicated the energy system is very dependent on the initial boundary conditions. In the case of ion plasmas associated with crop formations, the listing in Table 1 outlines the boundarycondition energies which, from the authors findings, appear to be involved in the organization of these complex vortex systems - selforganized through chaotic mechanisms. In a selforganized plasma the system is far from thermodynamic equilibrium, therefore, any slight change in any one of the seven energy components listed in Table 1 could produce a significant alteration in the structural makeup of the plasma system. In other words, a slight change in just one of these boundary conditions might lead to a drastic alteration in the geometric or nongeometric pattern produced at the crop surface. The complexity of influence from these boundary condition energies explains why no two crop formations ever have precisely the same geometric form.
If a crop formation occurs during, or subsequent to, a period of very violent weather the associated convective turbulence would tend to break up the organized plasma system into more localized, convective cells, thus producing crop formations with more irregular outlines - such as the roughly outlined, rectangular formation at the Blue Ball, MD, USA, sites. In fact it is important to note that both of the Maryland, 'super node expansion' formations were discovered less than 24 h after severe winds and thunderstorms passed through the area. These, and findings from other similar formations, led the authors to conjecture that weather with severe convective turbulence (energy component No. 4 in Table 1) breaks up the crop formation plasmas into smaller, but more energetic, convective cells of irregular outline. This more violent weather is characteristic of spring and summer conditions in the Midwest and Eastern United States. On the other hand, violent weather and tornadic winds are extremely rare in England and, consequently, the precisely formed, secondary instability products (Levengood 1958) take place under smooth, laminar flow conditions and are therefore more often transcribed into intricate, geometric design at the crop surface.
On rare occasions when a geometrically uniform crop formation has been produced, additional structures will be added within a period of one to several days - in the form of appendages, outer rings etc, adjacent to, but separate from, the initial formation. So far, these augmented structures have only been recorded at UK sampling sites. At one, a 1995 formation in barley at Beckhampton, England, the authors were fortunate to obtain normal upright control plants at sampling sites outside the initial formation before these control sites were (4 days later) incorporated into an added, outer flattened region. In Fig. 8A the aerial view displays a very uniform spiral formation (about 32 m diameter) formed May 28-29 1995 which, in turn, is surrounded by a 2.7 m wide path. In the aerial photograph this outer path is observed to have a different optical reflectance texture, when compared with the inner spiral complex or with the outer, standing plants. This effect is a result of a mixture of partially flattened and partially upright plants, typical of a region receiving very localized convective turbulence.
From the results shown in Fig. 6 it might be expected that the node length changes in the turbulent energy, outerring area would display greater node expansion when compared with the innerspiralring formation and, as shown in Fig. 8B this is precisely the case. The NL changes are as high as ÿ109% in the outer strip compared with a ÿ40% maximum in the plants within the initially formed (May 28-29), flattened spiral rings. If the situation within the initial, spiralringarea is examined, it is observed that a x 2 analysis demonstrates no significant difference between the distribution of node lengths in samples taken from the upright spiral rings compared with what would be expected by pure chance. In the alternate, flattened ring plant samples, the data at the lower part of Fig. 8B indicate significant node expansion. Because the sampling was conducted within 72 h after the formation appeared, it can be seen from Fig. 2 that less than 10% of the node changes in the downed rings can be attributed to gravitropic responses.
Conclusions
As data accumulate from the laboratory investigation of crop formation plants sampled at quite diverse geographic locations, it becomes apparent that the concept of organized ion plasmas as the guiding energetic force behind crop formations has a strong empirical foundation. The repeatedly documented lengthening and bending of crop formation plant stem nodes which have been sampled in formations just hours to a few days after their appearance in the fields, the frequent incidence of expulsion cavities, the fact that node length changes have been found to agree with the model of electromagnetic energy absorption described by the BeerLambert principle, all indicate the involvement of intense plasma energies, delivered very rapidly, during the crop formation process. In addition, a control study designed to evaluate two possible causative mechanisms for these observed plant anomalies - overfertilization and:or gravitropism - clearly revealed that neither factor contributes significantly to the documented plant alterations.
The further recognition that certain details of the flattened, intertwined crop 'lay' (such as the welldelineated sinusoidal 'ridge' pattern presented in Fig. 6A) can be explained as the result of two interacting plasma vortices with opposing rotation impacting the crop surface, adds significant credibility to the proposed hypothesis that thermodynamically unstable ionospheric, nonmeteorological, plasma systems are the causative agency.
Additional supporting evidence for this hypothesis has been obtained from rather unexpected sources, one example being a 'magnetic glaze' material discovered in a 1993 British formation. Detailed studies of this glaze, including scanning electron microscope (SEM) and energy dispersive spectroscopy (EDS), disclosed the presence of pure, iron oxides which had literally heat fused onto soil and plant leaves within very localized, plant swirls located within the boundaries of much greater circular regions of flattened plants.
Based on the physical, chemical and meteoritic nature of this iron oxide coating, the conclusion was reached (Levengood and Burke 1995) that clouds of meteoritic material from the August 1993 Perseid meteor showers had inter acted with a precrop formation, plasma complex.
 |
Fig. 8. Aerial view (A) of a 1995 spiral crop formation in barley at Beckhampton, England. The outer wide strip shown on the left in photograph (B), formed 5 days after the inner spiral. The values shown are percent node length change relative to the normal, upright controls. Node lengths from upright rings are not significantly different, whereas those from the downed rings are significant at PB0.05 (using standard t - test). |

Because meteoritic 'clusters' are known to be highly ionized and possess magnetic fields it was expected that, as they approached the large crop formation plasma system, they would experience a magnetic pinch effect (Rose and Clark 1960) which would tend to concentrate the meteoritic iron dust material into tight helical patterns and through mutual interaction of magnetic fields (boundary condition 6 in Table 1) they would be drawn into the large, precrop formation energy system where they would ultimately be melted and then deposited within the flattened crop area. Subsequent to the discovery of this meteoritic glaze in England in 1993, vigorous soil sampling at other crop formation sites, particularly in the USA, has revealed the presence of similar meteoritic residues.
These multiple findings continue to unveil, in both a quantitative and qualitative manner, the mechanics of interacting chaotic systems as a powerful tool for describing complex interactions within crop formation energy systems. Acknowledgements - The authors are especially grateful to George Reynolds for his dedicated fieldwork, particularly in implementing the 1997 Maryland control study, and to Barry Reynolds, Andy Thomas and the East Sussex field team and Peter Stammers and associates for their meticulous sampling efforts, as well as to artist Cheryl Gordon for her excellent assistance with the diagrams, and to British crop formation photographer, Steve Alexander, for sup plying both aerial and ground photographs of English crop formations.
References
Esau K (1967) Plant Anatomy. John Wiley and Sons, New York, p 689
Leopold AC (1964) Plant Growth and Development. McGrawHill, New York
Levengood WC (1958) Instability effects in vortex rings produced with liquids. Nature 181: 1680-1681
Levengood WC (1994) Anatomical anomalies in crop formation plants. Physiol Plant 92: 356-363
Levengood WC, Burke JA (1995) Semimolten meteoric iron associated with a crop formation. J Sci Exp 9: 191-199
Levitt J (1980) Response of Plants to Environmental Stresses, Vol II. Acad. Press, Inc. NY. ISBN 0124455026 (v.2)
Moore WJ (1962) Physical Chemistry. PrenticeHall, Englewood Cliffs, NJ, pp 820-821
Ortega JKE (1990) Governing equations for plant cell growth. Physiol Plant 79: 116-121
Peratt AL (1992) Physics of the Plasma Universe. SpringerVerlag, New York. ISBN 0387975756
Prandtl L, Tietjens OG (1957) Fundamentals of Hydro and Aeromechanics. Dover Pub., Inc., NY, pp 208-214
Rivest RL (1992) Vortex air rings. Science 356: 199-200
Rose DJ, Clark M Jr (1960) Plasmas and Controlled Fusion. MIT Press, Cambridge, John Wiley and Sons, NY
Salisbury FB, Ross C (1992) Plant Physiology. Wadsworth Pub. Co., Belmont, CA. ISBN 0534151620
Wilkins MB (1984) Advanced Plant Physiology. Pitman, London pp 170-174. ISBN 0273018531
Report a Crop Circle